






LncRNA HCP5參與卵巢早衰
卵巢早衰(POI)的遺傳病因至今已被證實,但lncRNAs在POI中的作用尚不清楚。那么,小編給大家介紹發表在“Nucleic Acids Research”上的文章“Long noncoding RNA HCP5 participates in premature ovarian insufficiency by transcriptionally regulating MSH5 and DNA damage repair via YB1”,讓大家詳細了解LncRNA HCP5在POI中的作用機制。

在本研究中,我們在生化性POI(bPOI)患者的顆粒細胞(GCs)中發現了一種低表達的lncRNA HCP5,它破壞了DNA損傷修復,促進了GCs的凋亡。在力學上,我們發現HCP5穩定了Nyb1和ILF2之間的相互作用,它可以介導YB1向GCs核的轉移。HCP5沉默影響了YB1在細胞核中的定位,降低了YB1與MSH5基因啟動子的結合,從而降低了MSH5的表達。
結果:
1.bPOI患者顆粒細胞中lncRNA HCP5表達下調
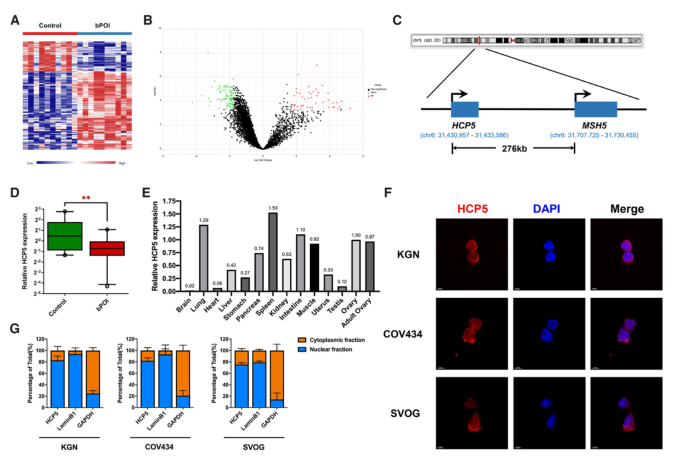
為了探討異常表達的lncRNAs與POI的關系,我們首先對bPOI患者和正常人的顆粒細胞進行了微陣列分析。我們發現159個lncRNA在患者和對照組之間有差異表達(圖1A,B)。在這些候選基因中,我們只考慮與POI相關基因共定位和共表達的lncRNA。lncRNA HCP5來自6號染色體MSH5基因上游276kb,是唯一一個POI基因共定位的lncRNA(圖1C)。定量RT-PCR分析證實,bPOI患者和對照女性的GCs中HCP5的表達存在差異(圖1D)。此外,我們的結果表明,HCP5在21周人胎兒的多個組織中廣泛表達,在卵巢中表達相對較高(圖1E)。采用熒光原位雜交(FISH)技術檢測HCP5的亞細胞定位。結果表明,HCP5主要位于細胞核內,尤其是在KGN、COV434和SVOG細胞的核膜周圍(圖1F),細胞分離結合qPCR分析進一步證實了這一點(圖1G)。
2.HCP5調節顆粒細胞MSH5的表達
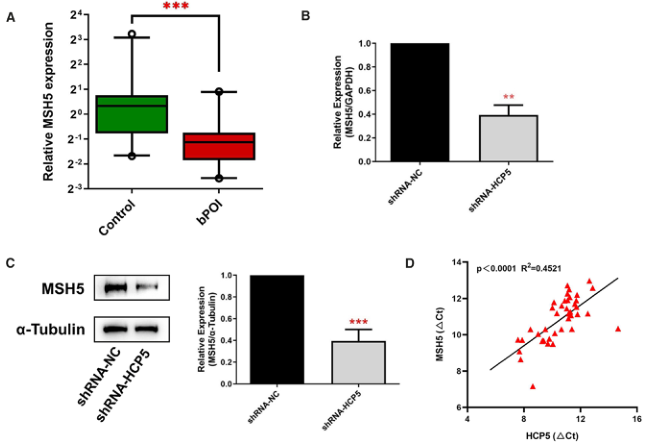
據報道,LncRNAs在其鄰近基因中起到順式調節作用。鑒于HCP5與MSH5的相鄰定位,我們提出HCP5可能參與MSH5的表達調控。RT-qPCR結果顯示bPOI女性GCs中MSH5較對照組下調(圖2A),這與HCP5的表達一致。此外,在KGN細胞中敲除HCP5導致MSH5在mRNA和蛋白質水平上的表達明顯降低(圖2B,C)。由于lncRNA-HCP5能在轉錄水平上調控MSH5的表達,故進一步研究HCP5是否與MSH5-mRNA在GCs中共同表達。我們檢測了GCs患者和正常人GCs中HCP5和MSH5的表達。如圖2D所示,lncRNA HCP5與MSH5 mRNA水平顯著相關。這些結果表明HCP5在調節MSH5表達中起著重要作用。
3.MSH5是DNA雙鏈斷裂(DSBs)修復過程中HCP5的靶點
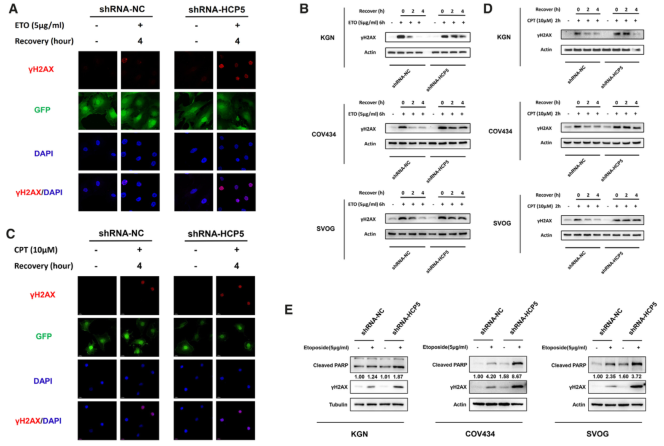
MSH5主要參與錯配修復,MSH4-MSH5雜二聚體在DSBs的同源重組(HR)修復中起重要作用。通過免疫熒光法發現DSBs的敏感指標γH2AX隨修復過程逐漸消失(圖3A)。western blot檢測γH2AX蛋白水平。結果表明,HCP5基因敲除明顯延長了ETO誘導的KGN、COV434和SVOG細胞DSB修復的時間(圖3B)。鑒于MSH5在DSBs的HR途徑中的重要作用,我們研究了HCP5基因敲除對喜樹堿(CPT)誘導的DNA損傷修復的影響。與ETO處理一致,γH2AX病灶被觀察到, 并隨著恢復逐漸消失(圖3C)。此外,沉默HCP5明顯延長了CPT誘導的KGN、COV434和SVOG細胞DSB修復的時間(圖3D)。此外,HCP5基因的敲除顯著增強了ETO誘導的細胞凋亡(圖3E)。
4.HCP5直接與YB1結合,是其核定位的關鍵
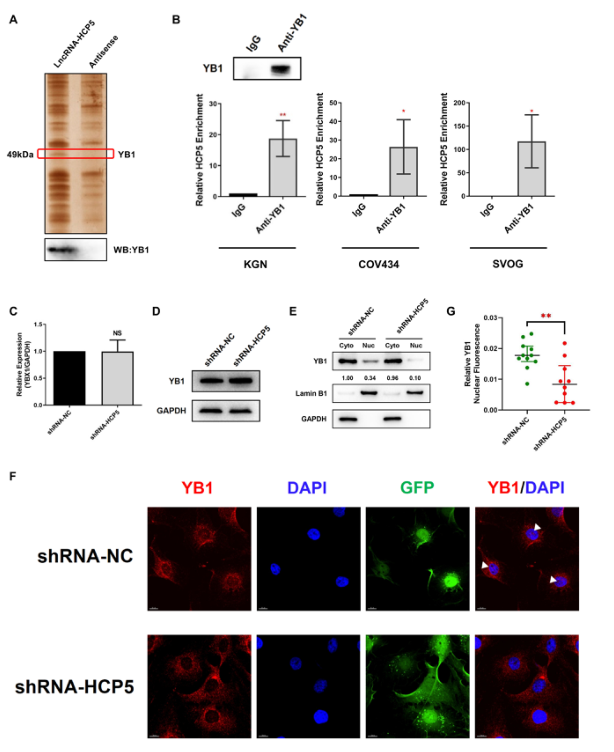
為了研究HCP5的蛋白伴侶,采用生物素標記的HCP5全長轉錄物及其反義轉錄物進行RNA下拉和MS聯合檢測。如圖4A所示,銀染后在50kDa處可見一條不同的HCP5結合蛋白帶。Western blot證實YB1在HCP5下拉蛋白中富集(圖4A)。由于RNA下拉實驗揭示了HCP5-YB1在體外的相互作用,我們采用RNA結合蛋白免疫沉淀法(RIP)檢測HCP5與YB1之間的內源性聯系。用抗YB1抗體進行RIP檢測,然后進行qRT PCR,證實了KGN、COV434和SVOG細胞中HCP5和YB1之間的直接相互作用(圖4B)。然后我們探討了HCP5對其結合蛋白YB1的調節作用。HCP5的敲除并沒有改變YB1在mRNA或蛋白質水平上的表達(圖4C,D),表明HCP5并不影響YB1在KGN細胞中的整體表達。考慮到YB1的活性主要取決于它在細胞核和細胞質之間的穿梭,進一步研究了HCP5是否參與了YB1的細胞定位。我們檢測了HCP5沉默后KGN細胞核質部分YB1的表達。Western blot顯示YB1存在于KGN細胞的細胞核和細胞質中;然而,HCP5沉默明顯減少了YB1在細胞核中的存在(圖4E)。免疫熒光分析表明YB1蛋白的核染色減少,排除了HCP5敲除時YB1的核仁定位(圖4F、G)。這些結果表明HCP5沉默阻止了YB1向細胞核的轉運。
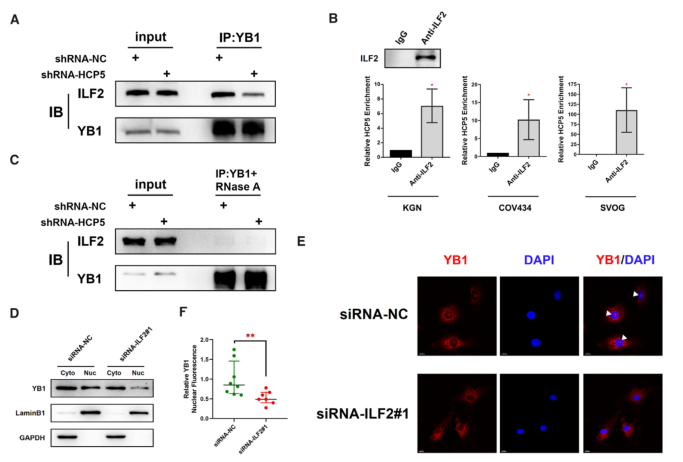
5.HCP5作為ILF2和YB1的RNA支架,對YB1定位到細胞核至關重要
在機制上,目前還不清楚HCP5是如何誘導YB1定位到細胞核的。考慮到ILF2可以與YB1相互作用并調節其核定位,我們采用共免疫沉淀的方法來評估HCP5對YB1 - ILF2復合物的影響。結果表明,HCP5的敲除降低了YB1抗體共沉淀的ILF2的相對量(圖5A)。考慮到HCP5在細胞核,特別是核膜中的定位,我們推測HCP5可能是YB1和ILF2之間的一種模塊化支架。因此,我們進行了RIP分析以確認HCP5和ILF2之間的相互作用。如預期,抗ILF2抗體在KGN、COV434和SVOG細胞中共沉淀HCP5(圖5B)。此外,我們觀察到在RNase A處理后YB1和ILF2之間的聯系減弱,這進一步驗證了RNA支架在YB1-ILF2相互作用中的重要作用(圖5C)。最后,ILF2沉默導致YB1的核定位顯著降低,這與HCP5基因敲除時的結果一致(圖5D-F)。因此,HCP5可以作為YB1和ILF2相互作用的RNA支架,并可能對YB1的核定位負責。
6.HCP5基因敲除降低了與MSH5啟動子的YB1結合并抑制了MSH5的轉錄激活
核YB1可以定位到啟動子或其他關鍵調控區的目標基因,并開始轉錄。我們進一步探討了HCP5敲除是否影響了MSH5啟動子對YB1的占用。染色質免疫沉淀(ChIP)分析和qPCR顯示YB1在MSH5啟動子上顯著富集(圖6A)。此外,HCP5的敲除顯著降低YB1與MSH5啟動子的結合(圖6B)。接著,我們研究了HCP5基因敲除對RNA PolⅡ與MSH5啟動子結合的影響。如圖6C所示,HCP5沉默導致RNA Pol II與MSH5啟動子的結合減少,這與HCP5 shRNA對MSH5 mRNA水平的抑制一致。綜上所述,HCP5通過促進YB1和RNA PolⅡ向MSH5啟動子的募集來調控MSH5的轉錄。
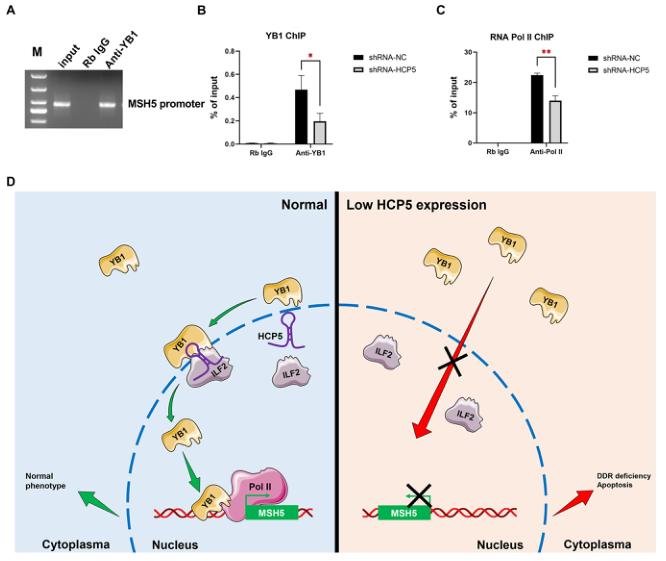
結論:我們闡明了LncRNA HCP5對人類POI的作用機制,即HCP5通過直接與YB1結合并調控其亞細胞定位來調節MSH5的表達和GCs的功能。本研究為POI的發病機制提供了一種新的表觀遺傳學機制。