






蛋白精氨酸甲基化修飾調控病毒感染后的MAVS激活
信號適配體MAVS在病毒感染后形成類似朊病毒的聚合體,以激活先天的抗病毒免疫反應。然而,MAVS的自發聚集可導致自身免疫性疾病。防止MAVS在靜止細胞中自發聚集的分子機制一直是個謎。在此,本研究發現,蛋白精氨酸甲基轉移酶9(PRMT9)直接針對MAVS,并催化MAVS在Arg41和Arg43處的精氨酸甲基化。在靜止狀態下,這種修飾抑制了MAVS的聚集和MAVS的自動激活。病毒感染后,PRMT9與線粒體解離,導致MAVS的聚集和激活。本研究揭示了MAVS上的一種翻譯后修飾形式,它可以使MAVS在生理條件下保持不活躍,以維持先天免疫的平衡性。本文于2022年8月發表在《Nature Communications》IF:17.694。
技術路線:

主要實驗結果:
1、PRMT9負調控RLRs誘導的IFN-β產生
為確定PRMTs在先天性抗病毒免疫中的潛在功能,將GFP-PRMT1-9質粒與DsRED2-Mito質粒一起轉染HEK293T細胞,然后進行模擬感染或感染SeV 8小時,并測量線粒體的定位。共聚焦顯微鏡顯示,PRMT1、PRMT2、PRMT3、PRMT4、PRMT5、PRMT6和PRMT8未能與線粒體共聚焦,且其定位不受SeV感染的影響。而PRMT7在HEK293T細胞中與線粒體共定位,不受SeV感染的影響,只有PRMT9在HEK293T細胞中與線粒體共定位,并在SeV感染后與線粒體分離(附圖1)。因此,選擇PRMT9用于后續實驗。

附圖1 PRMT9定位于線粒體
為進一步確認PRMT9的功能,通過CRISP/Cas9技術構建了兩個PRMT9敲除的RAW264.7細胞系(圖1a)。與siRNA敲除結果一致,在SeV感染或5′-pppRNA轉染后,PRMT9敲除的RAW264.7細胞中Ifnb1、Ifnα4和Cxcl10的mRNA表達水平上調(圖1b,c)。同樣,在RAW264.7細胞中敲除Prmt9也增加了VSV誘導的Ifnb1的表達(圖1d)。同時,在PRMT9敲除的RAW264.7細胞中,VSV的復制也減弱了(圖1d,e)。
在HEK293T細胞中轉染Flag-PRMT9,發現過表達Flag-PRMT9后,SeV或VSV感染后IFNB1 mRNA的表達明顯下降(圖1f,g)。同時,在過表達Flag-PRMT9的HEK293T細胞中,VSV的復制增加(圖1g)。熒光顯微鏡和流式細胞儀分析的結果表明,PRMT9在HEK293T細胞中的過表達促進了VSV-GFP的復制(圖1h)。總的來說,這些數據表明,PRMT9負向調節RLRs誘導的IFN-β信號,促進RNA病毒感染。
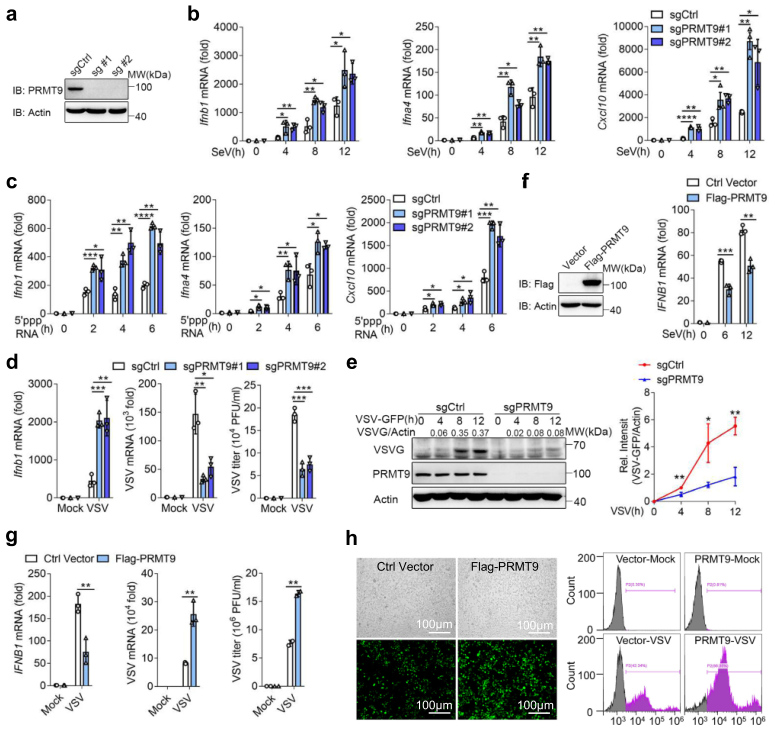
圖1 PRMT9負調控RLRs誘導的IFN-β產生
2、PRMT9缺陷增強RLRs觸發的IFN -β信號通路
為進一步研究PRMT9在調節病毒感染中的功能,將Prmt9fl/fl小鼠與Lyz2-Cre轉基因小鼠雜交,產生髓系特異性PRMT9敲除小鼠。從Prmt9fl/fl Lyz2-Cre(以下簡稱'Prmt9CKO')和Prmt9fl/fl小鼠(以下簡稱'Prmt9WT')制備初級腹膜巨噬細胞,然后用SeV、VSV和HSV-1感染或用5′-pppRNA刺激。與PRMT9的siRNA敲除或sgRNA敲除的觀察結果一致,用SeV感染Prmt9CKO腹腔巨噬細胞或用5'PPP RNA刺激導致Ifnb1、Ifnα4和Cxcl10 mRNA的表達與Prmt9WT巨噬細胞相比明顯增加(圖2a, b)。Prmt9CKO巨噬細胞的IFN-β分泌也比Prmt9WT巨噬細胞高(圖2a,b)。然而,Premt9WT巨噬細胞和Premt9CKO巨噬細胞在HSV-1感染誘導的I型IFNs和Cxcl10表達水平方面沒有觀察到差異(圖2c)。結果顯示,感染VSV后,Ifnb1 mRNA水平在Prmt9CKO巨噬細胞中增加,而VSV mRNA、VSV滴度和VSV蛋白在Prmt9CKO巨噬細胞中明顯下降(圖2d,e)。總之,可以得出一個結論:PRMT9對RLRs誘導的IFN-β信號傳導有負向調節作用。
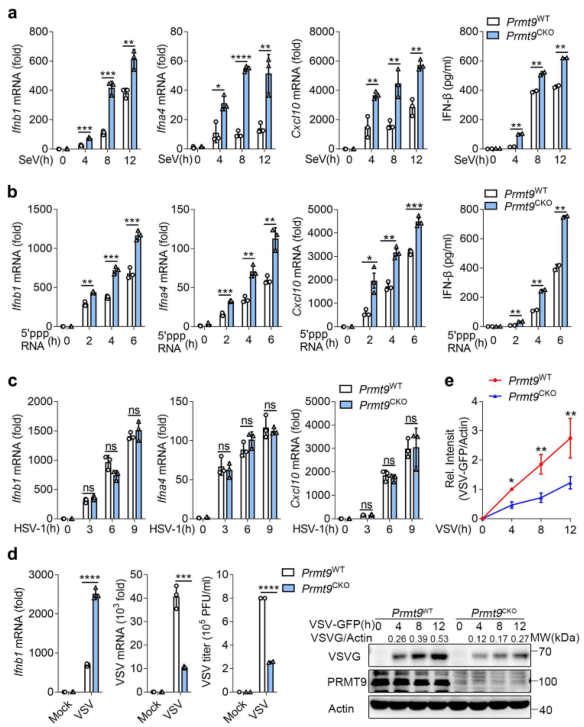
圖2原代腹腔巨噬細胞PRMT9缺陷增強RLRs觸發的IFN -β信號通路
3、在體內PRMT9抑制先天抗病毒響應
為確認PRMT9在RNA病毒感染中的生理功能,通過尾靜脈注射的方式讓Prmt9CKO小鼠感染VSV。如圖3a所示,感染VSV后,Ifnb1在Prmt9CKO小鼠肺部、肝臟和脾臟的表達水平明顯高于Prmt9WT小鼠的器官。此外,ELISA結果還顯示,VSV感染24小時后,Prmt9CKO小鼠血清中IFN-β蛋白的水平比Prmt9WT小鼠明顯增加(圖3b)。此外,VSV滴度的Plaque檢測和VSV mRNA的qRT-PCR分析證實,與Prmt9CKO小鼠相比,VSV在肺部、肝臟和脾臟的復制明顯減弱(圖3c, d)。此外,發現Prmt9CKO小鼠對VSV感染的敏感性低于Prmt9WT小鼠,但對HSV-1感染的敏感性不高(圖3e,f)。HE染色的結果顯示,Prmt9WT和Premt9CKO小鼠在沒有感染VSV的情況下,肺部沒有明顯的差異,與VSV感染后的Prmt9WT小鼠相比,PRMT9的缺乏減輕了炎癥細胞浸潤、組織水腫和肺部纖維化(圖3g)。總之,這些結果表明,PRMT9的缺失增強了體內對RNA病毒的抗病毒先天免疫反應。
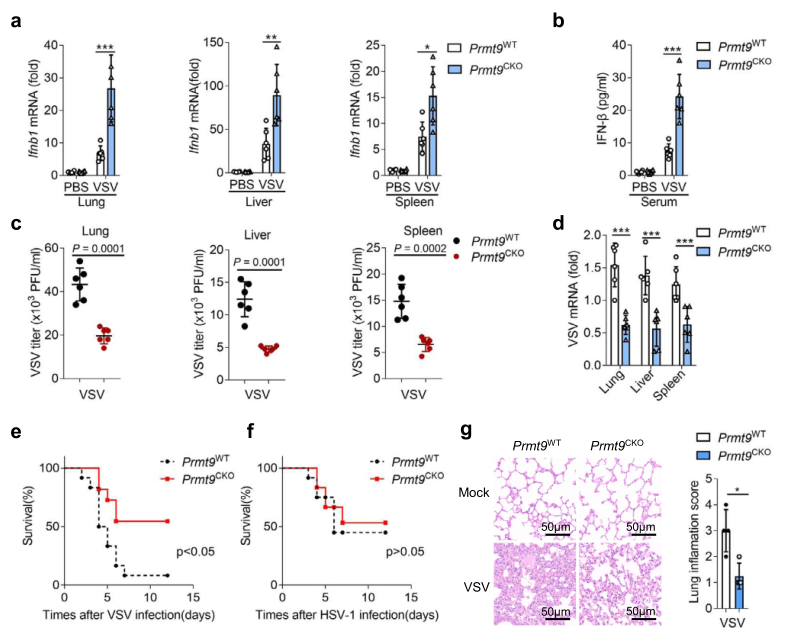
圖3 在體內PRMT9抑制先天抗病毒響應
4、病毒感染期間PRMT9負調控MAVS介導的信號通路
為研究PRMT9在調節MAVS介導的信號通路中的作用,將Flag-PRMT9質粒轉染到HEK293T細胞,然后用SeV感染。免疫印跡分析結果顯示,與空載體相比,轉染PRMT9表達質粒的HEK293T細胞中SeV誘導的TBK1、IRF3和IκBα的磷酸化水平下降(圖4a,b)。來自Prmt9WT和Premt9CKO小鼠的腹膜巨噬細胞,然后用SeV和HSV-1感染或用5'pp RNA刺激。發現,感染SeV或用5'pp-RNA刺激后,Prmt9CKO巨噬細胞中TBK1、IRF3和IκBα的磷酸化水平高于Prmt9WT巨噬細胞(圖4c, d)。相比之下,感染HSV-1后,TBK1、IRF3和IκBα的磷酸化在Prmt9CKO巨噬細胞中沒有表現出差異(圖4e,f)。據報道,在磷酸化后,IRF3將二聚體化并轉入細胞核。因此,從SeV感染的巨噬細胞中分離出細胞質和核的部分,測量IRF3的核轉位。發現在感染SeV后,Prmt9CKO巨噬細胞中IRF3向細胞核的轉移比Prmt9WT巨噬細胞的轉移要大(圖4g,h)。此外,共聚焦顯微鏡的結果顯示,PRMT9在HEK293T細胞中的過表達削弱了IRF3向細胞核的轉位(圖4i)。總之,這些數據表明,PRMT9通過抑制MAVS介導的信號傳導抑制IFN-β的產生和先天抗病毒免疫。
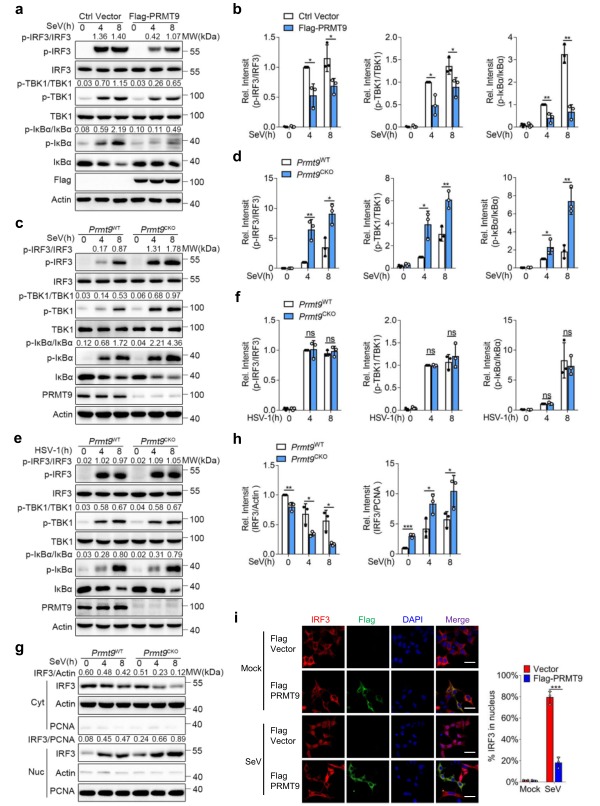
圖4 PRMT9負調控MAVS介導的信號通路
5、PRMT9靶向MAVS
為確定受PRMT9調控的目標,將與先天抗病毒免疫有關的主要信號蛋白的過表達質粒與PRMT9表達的質粒和IFN-β啟動子熒光素酶報告一起轉染到HEK293T細胞。發現由RIG-I、MDA5或MAVS介導的IFN-β啟動子熒光素酶報告的激活被PRMT9過表達所抑制(圖5a)。同時,由TBK1介導的IFN-β啟動子熒光素酶報告的激活不受PRMT9過表達的影響(圖5a)。cGAS/STING誘導的IFN-β啟動子的激活不受PRMT9過表達的影響(圖5a),這與數據顯示PRMT9不能調節對DNA病毒感染的抗病毒免疫反應相一致。進一步發現,HEK293T細胞中由RIG-I、MDA5或MAVS介導的IFNB1表達水平也受到PRMT9表達的抑制,而,由TBK1或IRF3介導的IFNB1表達水平在PRMT9過表達后沒有受到影響(圖5b)。這些數據表明,PRMT9可能以MAVS為目標,調節IFN-β信號和抗病毒反應。
為進一步驗證,將HA-cGAS、HA-RIG-I、HA-MAVS、HA-TBKI、Myc-MDA5和Myc-IRF3與GFP-PRMT9一起轉染到HEK293T細胞中,研究其相互作用。共同免疫沉淀和Western印跡分析的結果表明,PRMT9與MAVS相互作用,但與cGAS、RIG-I、MDA5、TBKI和IRF3沒有相互作用(圖5c)。用重組蛋白體外pull down的結果顯示PRMT9和MAVS之間有直接的相互作用(圖5d)。免疫熒光結果也顯示PRMT9和MAVS之間存在明顯的共定位(圖5e)。此外,發現病毒感染后,內源性PRMT9和MAVS之間的相互作用減弱了(圖5f)。總之,這些數據表明,PRMT9通過靶向線粒體上的MAVS抑制了抗病毒的先天免疫。
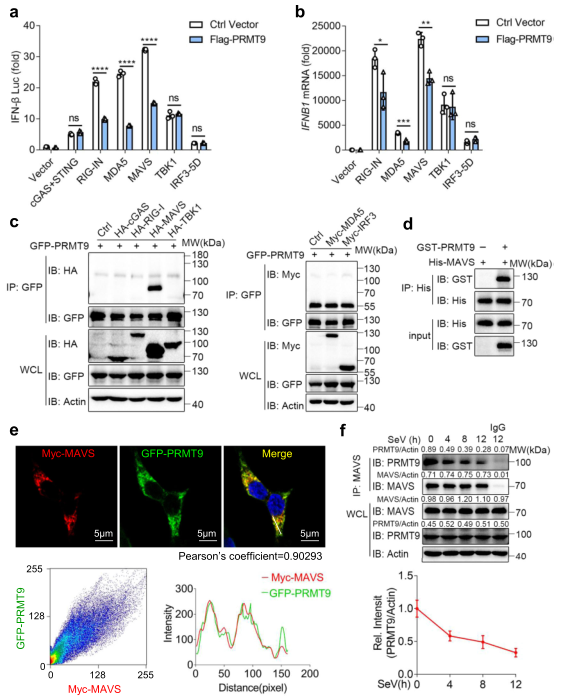
圖5 PRMT9靶向MAVS
6、PRMT9催化MAVS的R41和R43位點的精氨酸甲基化
接下來研究PRMT9是否能催化MAVS的精氨酸甲基化。首先將GFP-PRMT9表達質粒與HA-MAVS一起轉染到HEK293T細胞。Co-IP和WB結果顯示,PRMT9增強MAVS的SDMA水平(圖6a,左)。據報道,PRMT9能催化SAP14527的SDMA,因此,用HA-SAP145質粒做陽性對照(圖6a,右)。由于有三種類型的精氨酸甲基化是由精氨酸甲基轉移酶催化的,所以對MAVS的甲基化類型進行了研究。從小鼠中分離原發性腹膜巨噬細胞,用SeV感染。Co-IP和WB的結果顯示,在沒有SeV感染的MAVS中形成了MMA和SDMA,但沒有形成ADMA(圖6b)。值得注意的是,SeV感染后MMA和SDMA的形成逐漸減少(圖6b)。為進一步確定PRMT9的甲基化精氨酸功能,通過Co-IP和WB檢查內源性MAVS的SDMA。與PRMT9過表達的結果一致,在Prmt9CKO和Prmt9WT小鼠的腹腔巨噬細胞中敲除PRMT9大大減少了MAVS的甲基化(圖6c)。總的來說,這些數據表明PRMT9在RLRs途徑中調節MAVS的甲基化。
據報道,Gly260突變使PRMT9變成一種無催化活性的形式。為驗證PRMT9介導的MAVS甲基化是否取決于其甲基轉移酶的活性,將WT PRMT9或PRMT9突變體G260E與MAVS一起轉染到HEK293T細胞。結果表明,WT PRMT9可以增加MAVS的甲基化水平,而PRMT9的酶突變體G260E則失去了誘導MAVS甲基化的能力(圖6d)。SeV感染減少了PRMT9介導的MAVS甲基化(圖6d)。將WT PRMT9和PRMT9突變體G260E重新引入Prmt9CKO巨噬細胞并測量MAVS甲基化,發現在Prmt9CKO巨噬細胞中表達mPRMT9可以恢復MAVS甲基化,而表達mPRMT9突變體G260E則沒有這種效果(圖6e)。這些數據表明,PRMT9通過其甲基轉移酶活性催化了MAVS的甲基化。
為直接研究PRMT9催化MAVS的精氨酸甲基化,制備His-MAVS和GST-PRMT9的重組蛋白,并進行體外蛋白精氨酸甲基化試驗。發現在體外甲基化系統中,當PRMT9存在時,MAVS的甲基化程度增加(圖6f)。綜上所述,這些數據證明PRMT9促進了MAVS的精氨酸甲基化。
為確定MAVS中被PRMT9調節的潛在精氨酸殘基,用重組的MAVS和PRMT9蛋白進行體外甲基化試驗,然后用液相色譜-質譜(LC-MS)分析來識別被甲基化的精氨酸殘基。在檢測中,MAVS的R41和R43殘基被確定為潛在的甲基化部位(圖6g)。接下來,R41和R43精氨酸殘基被突變為賴氨酸,并檢查MAVS在MAVS KO HEK293T細胞(MAVS-/-)中的甲基化狀態。發現PRMT9可以促進MAVS(WT)的甲基化,而,MAVS(R41K)、MAVS(R43K)和MAVS(R41K,R43K)突變導致MAVS甲基化在PRMT9的存在下明顯減少(圖6h)。WB結果顯示,PRMT9可以促進MAVS(WT)的甲基化,而PRMT9介導的MAVS甲基化在MAVS(R41K)、MAVS(R43K)和MAVS(R41K, R43K)中大大降低(圖6i)。進一步,用S-[3H-Met]腺苷蛋氨酸作為甲基供體進行體外甲基化試驗。結果顯示,在重組PRMT9蛋白的存在下,His-MAVS的放射性大大增加(圖6j),而MAVS(R41K)、MAVS(R43K)和MAVS(R41K,R43K)的放射性水平與MAVS(WT)相比明顯下降(圖6j)。總之,這些數據表明,PRMT9催化了MAVS在R41和R43殘基上的精氨酸甲基化。
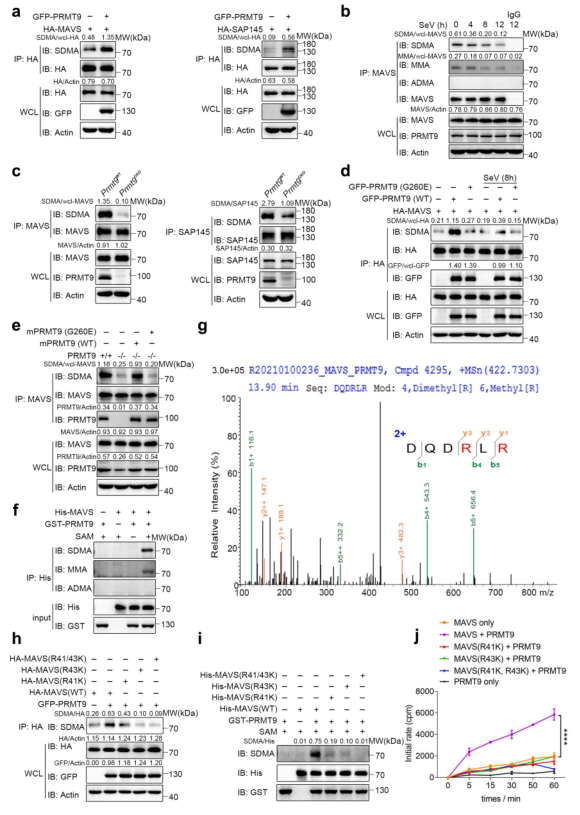
圖6 PRMT9催化MAVS的R41和R43位點的精氨酸甲基化
7、PRMT9介導的MAVS甲基化抑制MAVS聚集和激活
RNA病毒感染誘導MAVS迅速形成類似朊病毒的聚集物,導致I型IFN和其他炎癥細胞因子的強烈誘導。在本研究中,對PRMT9介導的MAVS甲基化是否抑制MAVS的聚集進行了調查。首先將GFP-PRMT9表達質粒或GFP-PRMT9(G260E)和HA-MAVS轉染到HEK293T細胞中,并通過半變性洗滌劑瓊脂糖電泳(SDD-AGE)測量MAVS的聚集。如圖7a,過表達MAVS可以促進MAVS聚集的形成。與PRMT9的抑制作用相一致,發現PRMT9過表達的HEK293T細胞中MAVS的聚集比轉染對照載體的細胞要少(圖7a)。值得注意的是,PRMT9(G260E)的過表達不能減少MAVS的聚集(圖7a)。進一步測量SeV感染誘導的MAVS聚集,該感染由Premt9CKO和Premt9WT小鼠的腹膜巨噬細胞制備。發現SeV感染可以誘導巨噬細胞中的MAVS聚集(圖7b),而PRMT9的缺乏大大增加了感染后Prmt9CKO巨噬細胞中MAVS聚集的形成(圖7b)。值得注意的是,即使沒有SeV感染,在Prmt9CKO巨噬細胞中也可以檢測到低水平的MAVS聚集(圖7b),表明PRMT9介導的MAVS甲基化可以阻止MAVS在正常條件下自主激活。
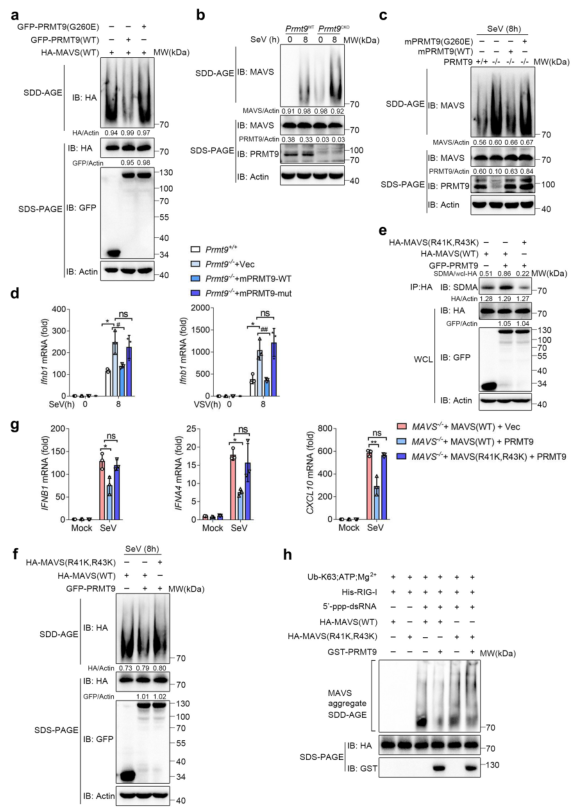
圖7 PRMT9介導的MAVS甲基化抑制MAVS聚集和激活
實驗方法:
CRISPR–Cas9構建PRMT9敲低小鼠,細胞和病毒培養,PRMT9和MAVS敲低細胞構建,質粒和轉染,RNA提取和質控,ELISA,熒光素酶報告基因實驗,Co-IP,IF,病毒感染和滴度實驗,蛋白表達和純化,組織病理染色,體外甲基化試驗,體外放射性實驗,體外MAVS的擴大和激活
參考文獻:
Bai X, Sui C, Liu F, Chen T, Zhang L, Zheng Y, Liu B, Gao C. The protein arginine methyltransferase PRMT9 attenuates MAVS activation through arginine methylation. Nat Commun. 2022 Aug 26;13(1):5016. doi: 10.1038/s41467-022-32628-y. PMID: 36028484