






單細胞和空間測序確定角質形成細胞和成纖維細胞放大銀屑病炎癥反應的過程
銀屑病是一種常見的皮膚慢性炎癥性疾病,其免疫發病機制尚不完全清楚。在這里,我們使用單細胞和空間RNA測序相結合的方法,證明了在沒有中性粒細胞蛋白酶的情況下,IL-36依賴性放大了IL-17A和TNF炎癥反應,這主要發生在銀屑病表皮的角質層上部。我們進一步展示了在銀屑病中,SFRP2+成纖維細胞的一個亞群通過轉變為促炎狀態而促進免疫網絡的放大。SFRP2+成纖維細胞通訊網絡涉及CCL13、CCL19和CXCL12的產生,通過配體-受體相互作用與其他空間上鄰近的細胞類型連接:CCR2+髓系細胞、CCR7+LAMP3+樹突狀細胞和CXCR4分別在CD8+Tc17細胞和角質形成細胞上表達。SFRP2+成纖維細胞也表達組織蛋白酶S,通過激活角質形成細胞中的IL-36G進一步放大炎癥反應。這些數據提供了對銀屑病發病機制的深入看法,這擴大了我們對關鍵細胞參與者的理解,包括炎性成纖維細胞及其細胞相互作用。
該研究于2023年6月發表在《Nature communications》,IF:16.6。
技術路線

1、對正常人和銀屑病患者皮膚單細胞的普查顯示了皮膚細胞群體的多樣性
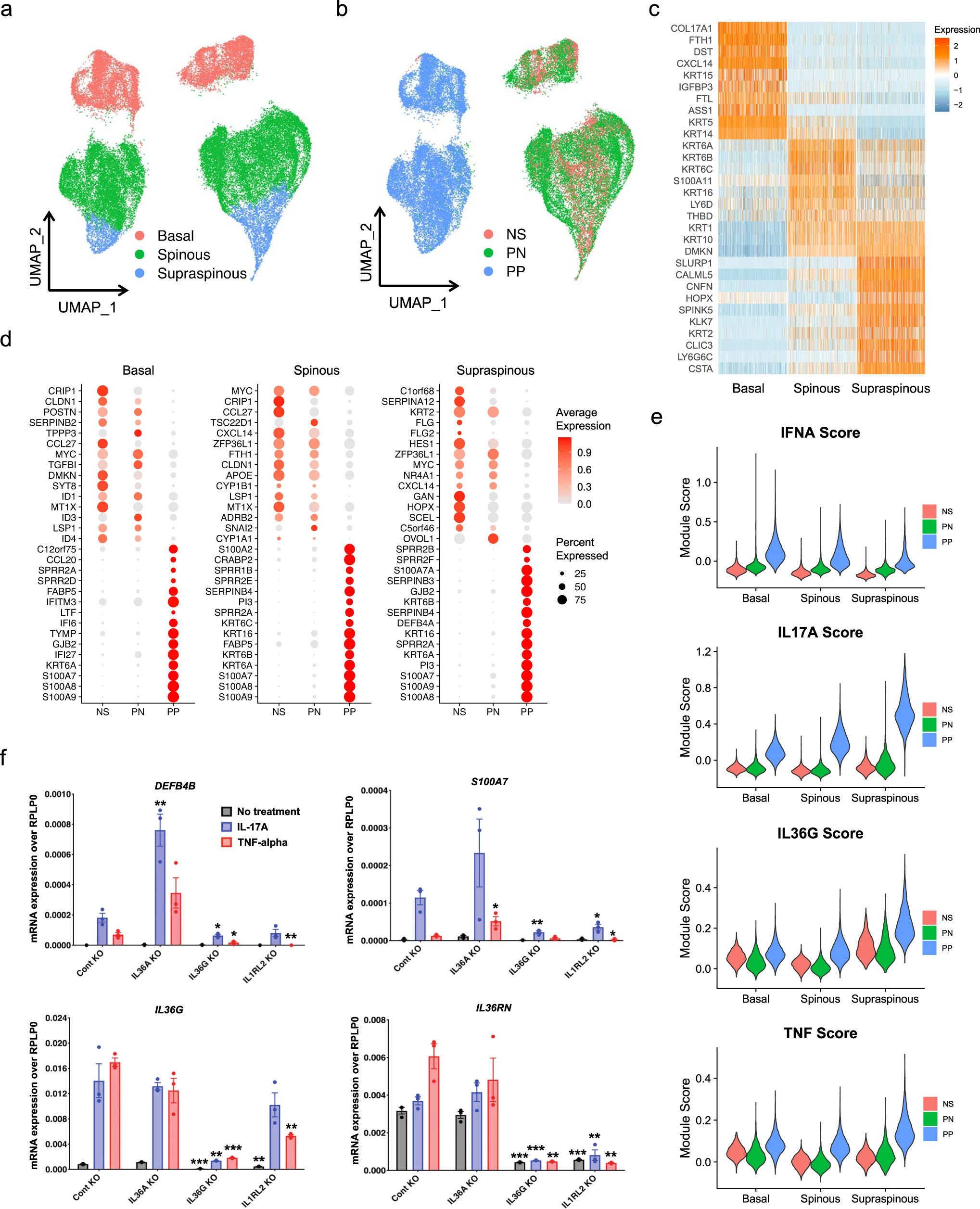
為了了解健康和銀屑病人皮膚的無偏細胞組成和細胞狀態,我們從8個健康人和14個銀屑病活檢標本中分離了皮膚細胞的單細胞懸液。我們還收集了來自同一批銀屑病患者的11個活檢標本中損傷周圍的皮膚細胞,總共得到了33個10X Chromium單細胞文庫(8個健康皮膚文庫(NS),11個銀屑病病變周圍皮膚文庫(PN)和14個銀屑病病變皮膚文庫(PP))。經過質量控制的銀屑病單細胞圖譜包括67,378個細胞,每個細胞檢測到的平均基因數為2421個,轉錄本數為10,953個(圖1a、b)。為了研究這些細胞的異質性,我們挑選可變基因,并使用統一的流形逼近和投影(UMAP)進行降維和R軟件包Seurat進行細胞聚類。通過將聚類標記基因與經典細胞類型定義的特征基因重疊,對聚類進行了驗證。我們鑒定出了10個主要的細胞類型,包括角質形成細胞(KC;KRT14、KRT1、DMKN)、黑素細胞(MLNC;DCT、TYRP1、PMEL)、汗腺細胞(ECG;PIP、DCD、MUCL1)、內皮細胞(EC;PECAM1、CDH5、CLDN5)、成纖維細胞(FB;DCN、COL1A1、COL1A2)、平滑肌細胞(SMC;ACTA2、TAGLN、MYL9)、神經細胞(Nerve;MPZ、PLP1、S100B)、T細胞(TC;CD3D、CD3E、TRAC)、髓系細胞(ML;CD74、HLADRA、HLA-DPB1)和肥大細胞(Mast;CPA3、TPSAB1、CTSG)(圖1c)。
在UMAP中,PP角質形成細胞與NS和PN角質形成細胞完全分離,這表明銀屑病角質形成細胞的基因轉錄組發生了根本性的改變(圖1b)。在獲得的細胞數量方面,最豐富的細胞類型是角質形成細胞,其次是成纖維細胞和T細胞。對每種細胞類型的疾病組成分析顯示,PP中免疫細胞和內皮細胞的比例增加,而汗腺細胞、神經細胞和黑素細胞的比例減少。這些結果表明,銀屑病特征包括明顯的角質形成細胞轉錄組變化和免疫細胞(包括T細胞和髓系細胞)的積累。
為了定位銀屑病皮膚中的各種細胞類型,我們對銀屑病皮膚進行了空間測序。我們從一個20微米厚的冰凍切片上收集RNA,并將其放置在一個含有空間條形碼捕獲寡核苷酸的55微米孔陣列上,用于轉錄組分析。在經過質量控制步驟后,我們檢測到987個空間定義的區域,每個區域平均檢測到2782個基因和11,779個轉錄本(圖1d)。我們通過將空間基因表達與單細胞RNA測序中主要細胞類型的基因表達進行反卷積,為每個區域標注了細胞類型組成。反卷積結果顯示在散點餅圖中,每個空間陣列中含有一個餅圖。K-means聚類顯示出六個具有不同平均細胞類型組成的聚類,代表了角質形成細胞、平滑肌細胞、成纖維細胞、汗腺細胞、內皮細胞、髓系細胞和T細胞的聚集體(圖1d)。空間測序中每個聚類的標記基因與通過單細胞RNA測序確定的細胞類型標記基因一致,表明注釋準確(圖1e)。角質形成細胞位于表皮中。髓系細胞、T細胞和內皮細胞位于靠近表皮的淺表真皮,而成纖維細胞、平滑肌細胞和汗腺細胞位于更深層的真皮(圖1d)。我們還觀察到一些空白區域,其中捕獲到的mRNA數量較少,代表真皮內無細胞區域。同時,我們對另外三個銀屑病和兩個健康皮膚活檢標本進行了空間測序,觀察到類似的細胞類型和空間細胞聚集。

2、銀屑病皮膚中不同的角質形成細胞分化狀態反映了銀屑病中不同的細胞因子反應
PP與NS和PN角質形成細胞的明顯分離表明在患有銀屑病的皮膚中存在重大的表皮功能變化(圖1b)。為了進一步描述這一點,我們對角質形成細胞進行了亞群聚類,并通過分化狀態對其進行了注釋。我們的分析在基底層(COL17A1、DST、KRT15)、棘層(KRT6A、KRT6B、KRT16)和表棘層(SLURP1、KLK7、KRT2)定義了三個不同的角質形成細胞分化狀態(圖2a-c)。我們對每個表皮分化狀態內的PP和NS角質形成細胞進行差異表達分析,并確定了在銀屑病皮膚的表棘層中不同表達的基因(DEGs)數量最多。我們繪制了每個分化狀態的前15個上調和下調基因,并發現PN角質形成細胞在所有三個表皮層中表現出介于NS和PP角質形成細胞之間的中間表達模式(圖2d)。S100A7、S100A8和S100A9在PP的所有三層中明顯過表達,與之前的銀屑病研究結果一致。有趣的是,I型干擾素誘導的基因IFITM3、IFI27和IFI6在PP上調基因中排名靠前,主要定位于銀屑病皮膚的基底表皮層。KRT6基因和KRT16在棘層和表棘層中排名靠前。值得注意的是,C1orf68,在以前的基因組關聯研究(GWAS)中被確定為銀屑病的靶基因,特異地在表棘層中表達(圖2d)。
接下來,我們使用Ingenuity通路分析(IPA)評估了每個表皮層內差異表達基因(DEGs)的富集通路和上游調控因子的細胞因子。這表明在銀屑病皮膚中,干擾素信號通路在基底層和棘層中是最重要的調控因子,而在表棘層中的作用較小。IL-17通路主要富集在表棘層中,而在棘層和基底層中富集較少。銀屑病皮膚中其他上游炎癥反應的富集包括OSM、TNF、IL-36、IL-6、IL-1、IL-18、IL-21、IL-22和IL-27。
為了追蹤這些細胞因子的來源,我們計算了每種細胞類型在不同疾病狀態(NS、PN、PP)下這些基因的平均表達水平。這些炎癥介質的大多數由特定的細胞類型表達,例如,角質形成細胞表達IL18和IL36G;T細胞表達IL17A、TNF、IL21和IL22;髓系細胞表達IFNB1、IL1A、IL1B和IL27。值得注意的是,與PN和NS相比,IL36G、IL17A、IL21、IL22和IL27在相應的細胞類型中在PP中的表達最高。為了驗證這些關鍵上游調控因子,我們計算了通過單個促炎細胞因子(包括I型干擾素(IFN-α)、IL-17A、IL-36γ和TNF)刺激培養的角質形成細胞中誘導基因的模塊得分(圖2e)。與DEGs的富集分析一致,IFN-α得分在所有三個層次中從NS逐漸增加到PN再到PP,在基底層得分最高,棘層略有下降,表棘層進一步下降。IL-17A得分在NS和PN中在所有三個層次上相似,但在PP角質形成細胞中從基底層到棘層再到表棘層有顯著增加。類似地,在PP表皮中,IL-36γ得分在表棘層中比基底層和棘層更高。為了驗證這些反應的空間特異性,我們計算了角質形成細胞亞型模塊得分以及IL-17A和IL-36γ模塊得分。與scRNA-seq結果一致,IL-17A和IL-36γ模塊得分與表棘層模塊得分最高相關,而與棘層或基底層模塊得分的相關性較低。這些結果表明,在銀屑病中,不同的表皮組織受促炎細胞因子的影響程度不同。
為了研究IL-36在激活IL-17和TNF反應中的作用,我們使用CRISPR-Cas9在角質形成細胞中敲除(KO)IL36A、IL36G和IL1RL2(IL-36R),并測量IL-17和TNF誘導的四個抗菌和促炎基因的表達:DEFB4、S100A7、IL36G和IL36RN(圖2f)。敲除IL36G或IL1RL2都導致角質形成細胞中這四個基因的顯著減少,表明在中性粒細胞蛋白酶不存在的情況下,IL-36G和IL-36R直接增強表皮中IL17A和TNF的反應(圖2f)。值得注意的是,與之形成鮮明對比,從抗菌基因DEFB4和S100A7的mRNA表達來看,IL36A KO導致增強IL-17A和TNF的反應(圖2f)。
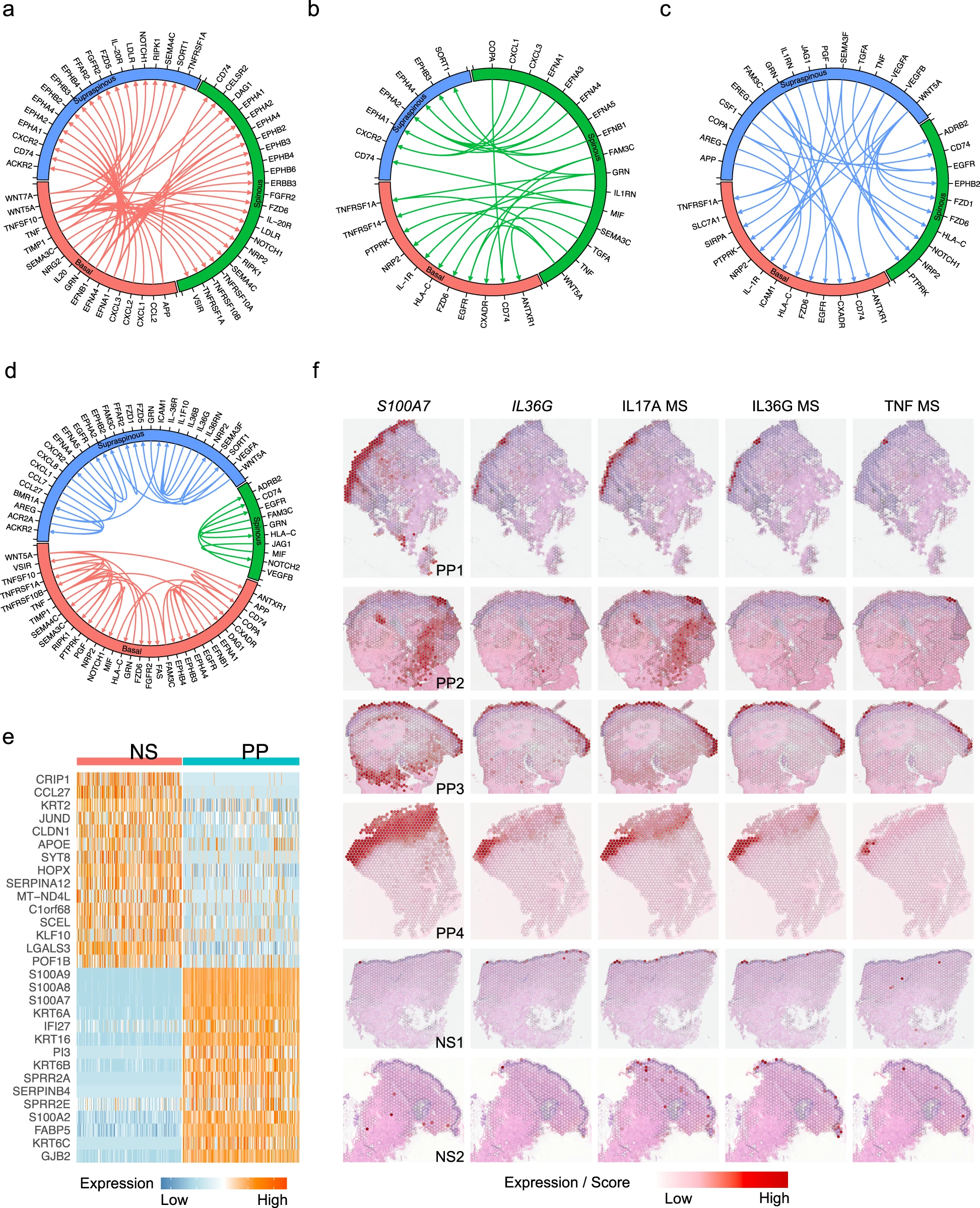
鑒于角質形成細胞在銀屑病炎癥反應中的重要性,我們試圖確定銀屑病皮膚中三個表皮層之間的配體-受體相互作用。為此,我們對所有NS角質形成細胞亞型和所有PP角質形成細胞亞型運行了CellPhoneDB,并分析了在PP中與NS相比具有較高相互作用得分的配對。三個層次中最豐富的配體-受體對出現在ephrin配體家族和其受體之間,這在表皮內穩態中起到關鍵的信號傳導作用(圖3a-c)。Notch信號通路在角質形成細胞層之間也是顯著的,并且已經報道在表皮生長和分化中發揮重要功能。有趣的是,IL-36信號僅在銀屑病患者的表棘層觀察到,與scRNA-seq和空間-seq的表達數據一致,它通過誘導表棘層角質形成細胞產生的趨化因子CXCL1和CXCL8進一步促進白細胞浸潤,尤其是中性粒細胞的浸潤到表皮中(圖3d)。
為了確定三種角質形成細胞亞型的空間位置,我們試圖通過scRNA-seq分析中定義的亞型表達特征來解析spatial-seq中的角質形成細胞斑點。預測得分準確反映了表皮三個層次的空間位置,棘上角質形成細胞位于最外層,棘狀角質形成細胞位于中間層,基底層角質形成細胞位于最內層。我們對PP角質形成細胞斑點和NS角質形成細胞斑點進行差異表達分析,并得到了與scRNA-seq相似的一組DEGs(圖3e)。兩個代表性基因S100A7和IL36G的空間定位圖顯示了它們在PP中的上調表達和在表皮中的特定位置(圖3f),這一結果也得到了scRNA-seq數據集的驗證。值得注意的是,IL36G主要在被預測為表棘層角質形成細胞的斑點中被檢測到,與我們的scRNA-seq分析中觀察到的細胞因子反應結果一致。體外培養的銀屑病皮膚活檢標本中遷移的角質形成細胞的scrna-seq檢測也檢測到了IL36G。我們還在spatial-seq樣本中繪制了IL-17、IL-36和TNF模塊得分,并驗證了IL36G和S100A7表達與這些細胞因子反應的共定位(圖3f)。
UMAP分析將NS和PP角質形成細胞分開,顯示了在健康和疾病條件下存在不同的角質形成細胞分化途徑(圖2b)。為了表征這些分化途徑,我們分別對NS和PP角質形成細胞進行了Monocle的擬時序分析。Monocle擬時序分析將細胞排列成一個線性軌跡,其方向沿著從基底層到棘層再到表棘層的方向排列,這對于NS和PP角質形成細胞都適用。為了確定推動分化的潛在細胞因子,我們將可變基因沿著擬時序分成五個表達模式,分別針對NS和PP軌跡。接下來,我們使用IPA推斷了每個表達模式中基因的上游調控因子。對于每個上游調控因子,我們使用五種表達模式的所有目標基因計算模塊得分。然后,我們計算了每個上游調控因子的模塊得分與Monocle分析定義的擬時序之間的相關性。我們發現,只有OSM和IL-4模塊得分與NS角質形成細胞的擬時序呈正相關,而包括IL-22、IL-1β、IL-17A、IL-13和TNF在內的多種細胞因子的模塊得分與PP角質形成細胞的擬時序高度相關。為了驗證驅動PP角質形成細胞分化的關鍵細胞因子,我們使用由單個細胞因子(包括IL-1β、IL-17A、IL-13和TNF)刺激培養的角質形成細胞中誘導的基因計算了模塊得分。這四種細胞因子的模塊得分與PP角質形成細胞的擬時序高度相關,與IPA推斷的結果一致。這些結果表明,在銀屑病中,角質形成細胞的分化受到多種細胞因子的局部產生的驅動,尤其是IL-1β、IL-22、IL-17A、IL-13和TNF。
角質形成細胞的過度增殖是銀屑病的一個顯著特征。為了解決角質形成細胞的增殖問題,我們研究了NS、PN和PP之間的細胞周期效應,通過添加被排除在我們最初評估之外的增殖角質形成細胞,以便于分析細胞的異質性。預期地,與NS或PN樣本相比,來源于PP樣本的增殖角質形成細胞超過50%。我們在spatial-seq樣本中繪制了細胞周期得分,發現表皮的增殖效應最強,特別是在表皮的基底層。我們還發現,在PP樣本中存在著比NS樣本更厚的增殖角質形成細胞層,從而證實了我們的spatial-seq數據中PP的高增殖表型。
3、SFRP2+成纖維細胞在銀屑病炎癥中的作用與其促纖維化作用不同
考慮到根據scRNA-seq數據定義的銀屑病病變中第二常見的細胞類型是成纖維細胞。我們將所有成纖維細胞分成了12個亞群(圖4a-c)。基于先前發表的皮膚成纖維細胞標記基因,我們將亞群注釋為四個亞型:SFRP2+、COL11A1+、SFRP4+和RAMP1+成纖維細胞(圖4b)。我們通過免疫組織化學(IHC)在銀屑病皮損中確認了主要成纖維細胞亞型的存在。SFRP2+成纖維細胞是皮膚中最大的成纖維細胞亞群,與先前的研究結果一致。盡管SFRP2+成纖維細胞包含來自PP、PN和NS的細胞,值得注意的是,使用UMAP進行降維時,PP成纖維細胞主要與PN和NS成纖維細胞分開(圖4c)。為了確定推動這種在銀屑病成纖維細胞中的轉變的機制,我們確定了PP和PN成纖維細胞之間的差異表達基因,并使用IPA進行富集分析。激活的上游調控因子分析確定了I型干擾素、干擾素-γ和TNF作為促進SFRP2+成纖維細胞亞群中銀屑病相關變化的top級介導因子(圖4d)。有趣的是,通過比較PP和PN細胞,我們發現這些上游調控因子在三個角質形成細胞亞型中也被激活,這表明這些是促使成纖維細胞和角質形成細胞從PN到PP轉變的共同驅動因子。
在SFRP2+成纖維細胞中,有四個亞群主要由PP成纖維細胞組成,分別是亞群3、0、6和2(圖4c,e)。有趣的是,差異表達分析顯示PP SFRP2+成纖維細胞存在顯著的異質性,亞群3標記物中包括膠原基因(例如COL1A1和COL3A1),而亞群6標記物中則包括一組促炎細胞因子(例如CCL19、IL33、IL34)以及蛋白酶CTSS(圖4f)。這些趨化因子和蛋白酶在上皮表皮層和真皮乳頭尖部的成纖維細胞中表達最為明顯,這是通過空間-seq映射和免疫熒光(圖4g)揭示的。對成纖維細胞亞群3和亞群6標記基因的上游調控因子分析顯示,關鍵的銀屑病相關細胞因子IFN-γ、TNF、IL-1β、OSM和IL-17A的激活z分數增加最多,而與纖維化相關的IL-5、EDN1和IL-4的激活z分數降低(圖4h)。綜上所述,這些結果揭示了PP SFRP2+成纖維細胞的異質性,包括膠原基質成分的產生者以及細胞因子誘導的非基質產生的促炎區域。

4、CD8+Tc17是銀屑病皮膚中IL17A的主要來源
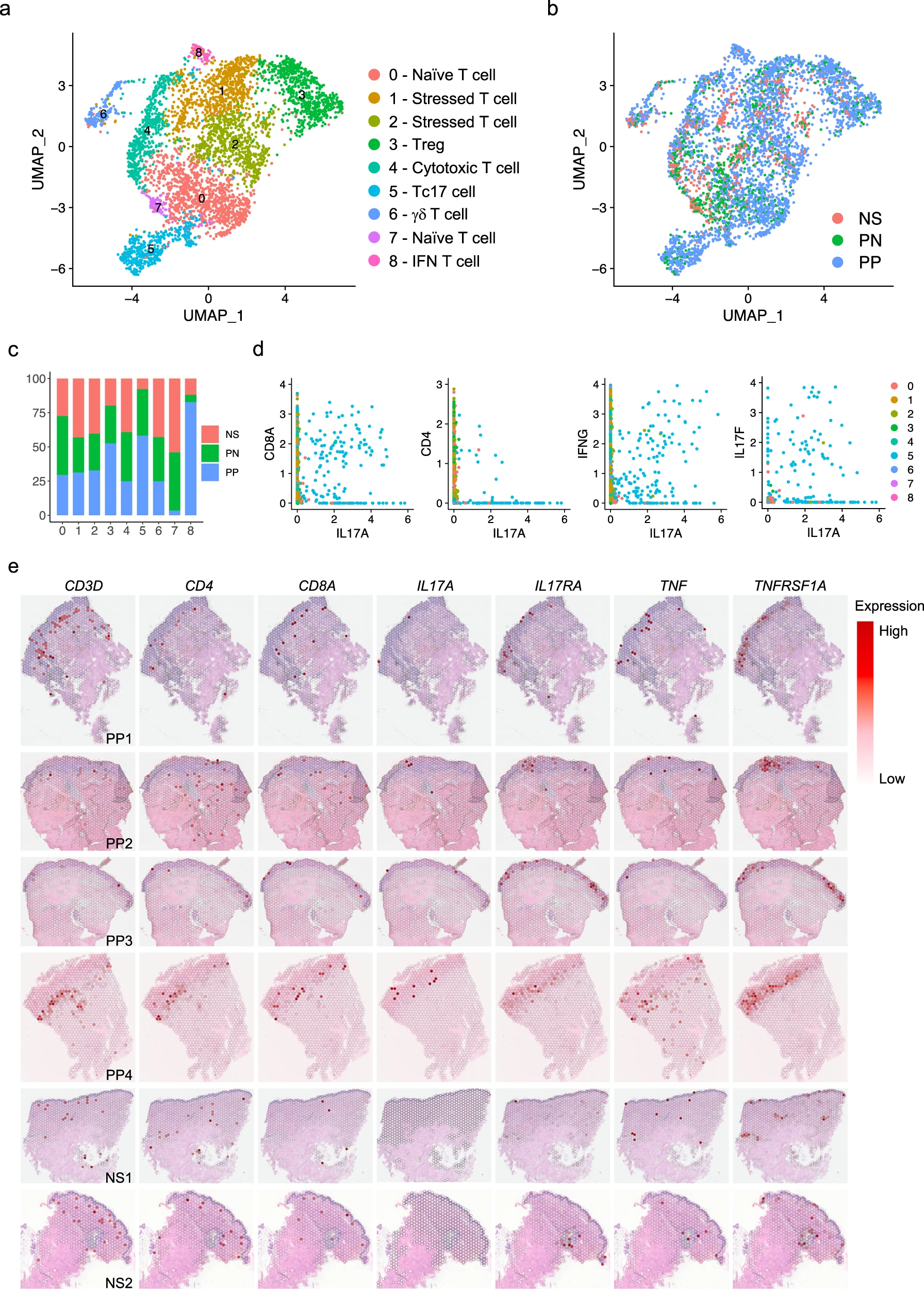
長期以來,產生IL-17的T細胞一直被認為是銀屑病發病機制的核心。為了定義T細胞亞型在銀屑病中的作用,我們對T細胞進行了亞群聚類,并使用經典的T細胞譜系標記對亞群進行了注釋(圖5a、b)。我們在正常皮膚(NS)、健康對照皮膚(PN)和銀屑病皮膚(PP)中鑒定出了七種T細胞亞型:原始T細胞(亞群0和7;RGCC、CCR7、IL7R)、應激T細胞(亞群1和2;DNAJB1、HSPA1A、HSPH1)、調節性T細胞(Tregs)(亞群3;FOXP3、TIGIT、BAFT)、細胞毒性T細胞(亞群4;CCL5、NKG7、GZMK)、分泌IL17的CD8 T細胞(Tc17細胞)(亞群5;IL17A、IL17F、CCL20)、γδT細胞(亞群6;TRDC、FCER1G、TYROBP)和IFN T細胞(亞群8;IFI6、MX1、IFIT3)。值得注意的是,大部分T細胞來源于銀屑病皮膚樣本,與已知T細胞在銀屑病皮膚中的大量滲入一致(圖5b)。對每個亞群的疾病組成分析顯示,在銀屑病中,Tregs、Tc17細胞和IFN T細胞的比例增加(圖5c)。與角質細胞的發現一致,PP中呈現出IFN特征的T細胞比例增加,表明在銀屑病皮膚中,強烈的IFN反應不僅影響角質細胞,還影響其他細胞類型。我們檢查了之前報道與銀屑病有關的細胞因子的表達情況,發現IL17A、IL17F、IL26、CCL20、CXCL13、IL22和IL23R專門由Tc17細胞表達。傳統上,認為產生IL-17的T細胞主要是CD4+ T細胞,然而最近的觀察為銀屑病中產生IL-17的CD8+ T細胞提供了越來越多的證據。從皮膚中鑒定到CD4+和CD8+ T細胞使得我們能夠直接比較這些T細胞亞群及其在銀屑病皮膚中對IL-17反應的貢獻。在這里,我們的數據表明CD8+ T細胞是銀屑病中IL17A的主要來源。為了進一步表征這一點,我們檢查了IL17A與CD4、CD8A和IFNG的共表達情況,發現IL17A表達細胞與CD8A和IFNG表達細胞呈正相關,但很少觀察到CD4+ IL17A+細胞(圖5d)。此外,我們的數據表明,IL17A產生的T細胞數量比IL17F產生的多,盡管大部分細胞同時表達這兩個因子(圖5f)。我們將T細胞標記基因映射到空間測序樣本中,并揭示出盡管在表皮中鮮有檢測到IL-17或TNF產生的T細胞,大部分T細胞在皮膚真皮表皮交界處檢測到(圖5e),這一結果在免疫熒光染色中得到了驗證。有趣的是,我們發現IL-17受體基因(IL17RA)和TNF受體基因(TNFRSF1A)在PP樣本的表皮中高表達(圖5e),與在角質細胞中發現的局部細胞因子反應結果一致。
5、單細胞RNA測序確定銀屑病患者獨特的髓系亞群
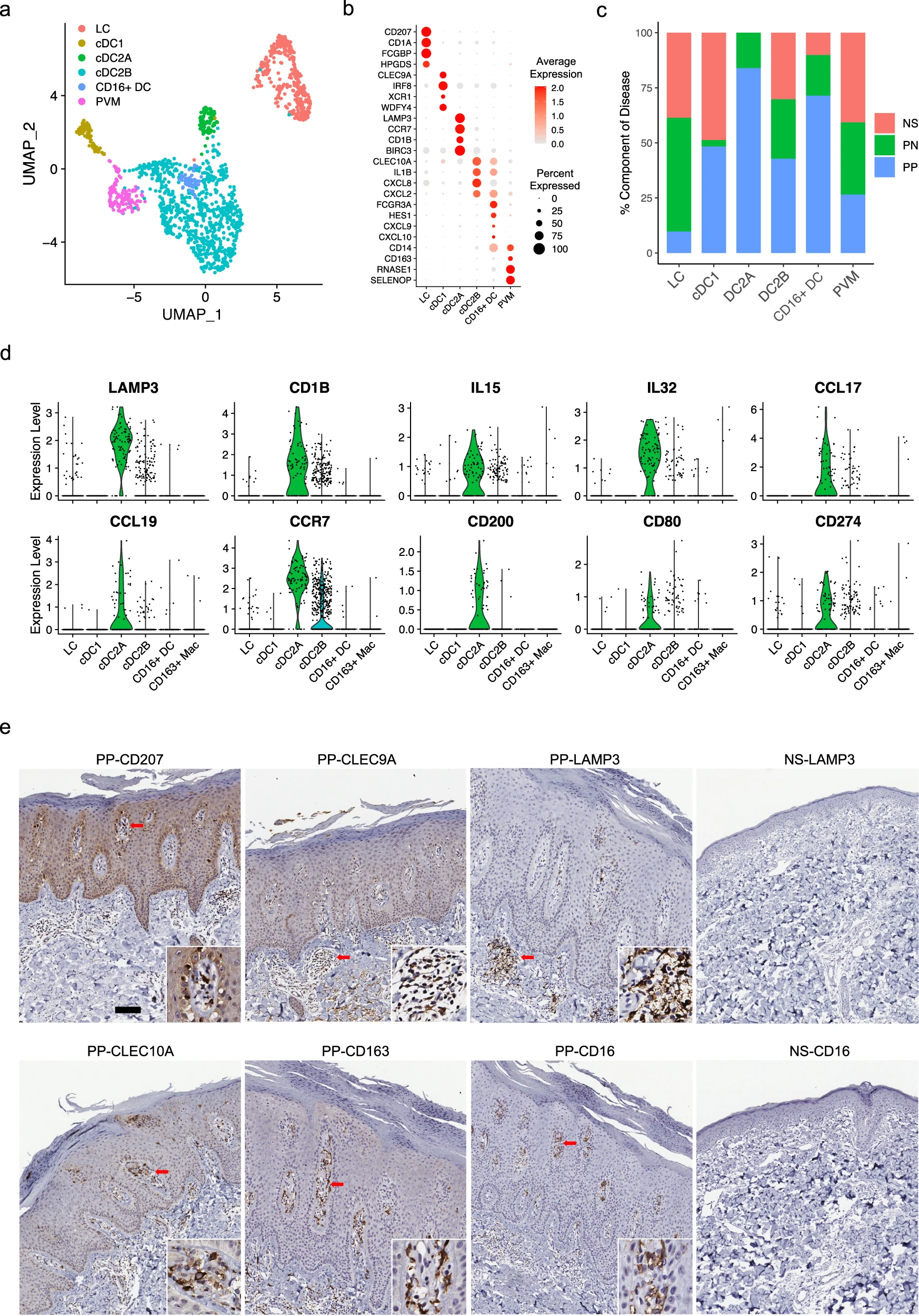
為了檢查銀屑病皮膚中髓系細胞的細胞異質性,我們對髓系細胞進行了亞群聚類,并注釋了六個髓系亞群:朗格漢斯細胞(LC;CD207和CD1A)、經典1型樹突狀細胞(cDC1;CLEC9A和IRF8)、經典2型樹突狀細胞A亞群(cDC2A;LAMP3和CD1B)、經典2型樹突狀細胞B亞群(cDC2B;CLEC10A和IL1B)、CD16+樹突狀細胞(CD16+ DC;FCGR3A和HES1)和周圍血管巨噬細胞(PVM;CD163和SELENOP)(圖6a、b)。對每個亞型的疾病組成分析顯示,在PP中LC的比例減少(圖6c)。先前的研究表明在銀屑病皮膚中LC的遷移和功能受損,我們的分析結果與此一致,顯示PP中LC的數量減少。值得注意的是,cDC2A僅來源于PN和PP皮膚,其中PP皮膚中的數量更多。先前的研究報告顯示IL15和IL32在銀屑病中表達增加,這兩種細胞因子都作為促炎介質,并且主要由cDC2A髓系細胞表達(圖6d)。我們還發現CCL17、CCL19及其受體CCR7主要由cDC2A表達(圖6d)。抑制CCL19/CCR7軸在病損皮膚中被報道為TNF阻斷誘導的銀屑病患者臨床緩解的關鍵事件。此外,cDC2A高表達共刺激分子CD200、CD80和CD274,表明這些細胞可能在銀屑病皮膚中引導和推動T細胞活性和擴增(圖6d)。CD16+ DC主要來源于PP樣本(圖6c)。CD16+ DC具有與cDC2B類似的表達模式,包括CLEC10A、IL1B和CXCL2,但也表達FCGR3A(CD16)、CD14和HES1,這表明它們可能起源于循環CD14+ CD16+單核細胞(圖6b)。值得注意的是,我們發現CXCL9和CXCL10主要由CD16+ DC表達,這兩種趨化因子已被證明通過STAT1、STAT4和STAT5激活驅動效應性Th1/Th17細胞極化(圖6b)。這些數據在銀屑病和正常皮膚中通過免疫組織化學染色得到驗證,并顯示了這些亞群在銀屑病皮膚中的不同定位,CLEC9A和LAMP3主要定位于真皮中的淋巴樣結構中,CLEC10A和CD16主要位于真皮乳頭的頂部(圖6e)。在正常皮膚中找不到LAMP3+和CD16+細胞。PVM位于真皮乳頭的周血管位置,而LC主要位于真皮乳頭的真皮-表皮交界處(圖6e)。
6、單細胞RNA測序突出銀屑病相關遺傳風險變異的細胞類型特異性
為了確定與銀屑病相關的遺傳變異相關的基因及其細胞位置,我們檢查了GWAS在銀屑病中檢測到的65個單核苷酸多態性(SNP)位點。使用人體皮膚的表達定量性狀位點(eQTL)數據庫,共鑒定了與這些SNP相關的46個基因。為了增加與SNP位點相關的基因數量,我們還考慮了人類基因組中與SNP位點最近的基因,共得到65個相關基因。在去除低表達基因后,我們得到27個與eQTL相關的基因和57個最近基因。我們計算了這些基因在NS、PN和PP中每個細胞類型中的平均表達,并確定了表達每個基因的細胞類型。值得注意的結果包括C1orf68基因在角質細胞中的主要表達,且從NS到PN再到PP呈遞減趨勢。IL23R基因在PN和PP的T細胞中主要表達,已被證明促進銀屑病中的IL-17反應。這些結果為功能研究提供了寶貴的資源,旨在了解與銀屑病相關的遺傳位點及其關鍵細胞群體的機制。
7、配體-受體分析顯示銀屑病的細胞類型特異性網絡
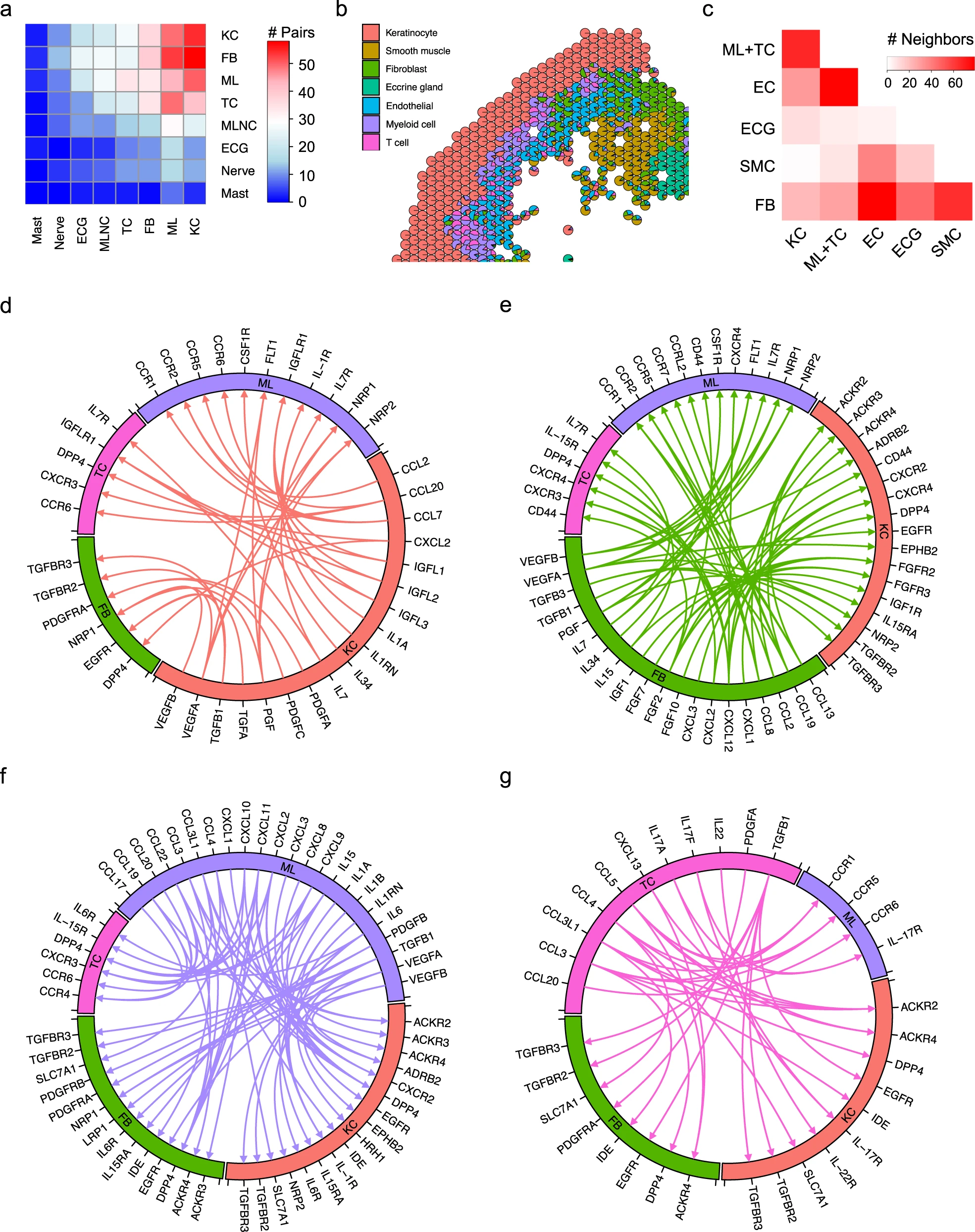
鑒于觀察到與健康皮膚相比,銀屑病皮膚中的細胞類型和亞型組成發生了變化,我們分析了隨著銀屑病的發展細胞間的相互通訊如何改變。為此,我們首先進一步對內皮細胞、平滑肌細胞和黑素細胞進行了亞群聚,并分別使用CellPhoneDB在NS特定的亞群聚和PP特定的亞群聚上進行配體-受體分析。為評估銀屑病中發生的變化,我們分析了在PP中與NS相比具有更高相互作用分數的配體-受體對。在銀屑病皮膚中,配體-受體對變化最多的細胞類型有四種:角質細胞、成纖維細胞、髓樣細胞和T細胞(圖7a)。我們認為空間位置上的鄰近性將促進細胞間的通訊。為此,我們統計了每對細胞類型的相鄰斑點數,如散點-餅圖所示(圖7b)。結果表明,角質細胞具有最多的免疫細胞鄰居(T細胞和髓樣細胞),成纖維細胞分布在許多其他細胞類型中,內皮細胞與免疫細胞相鄰(圖7c)。通過匯總配體-受體對變化的數量和空間鄰近性分析,我們將分析重點放在角質細胞、成纖維細胞、髓樣細胞和T細胞之間的相互作用上。
值得注意的是,角質細胞預測的相互作用包括CCL2和CCL7與髓樣細胞中的CCR2的相互作用,CCL20與髓樣細胞和T細胞中的CCR6和IL7與IL7R的相互作用,以及類胰島素生長因子家族配體(IGFL1、IGFL2、IGFL3)與其受體(IGFLR1)在T細胞上的相互作用(圖7d),這表明角質細胞在調控銀屑病中的免疫反應中起著重要作用。此外,角質細胞表達了血小板源性生長因子(PDGFA、PDGFC)、轉化生長因子(TGFB1)、表皮生長因子(TGFA)和血管內皮生長因子(VEGFA、VEGFB),這些因子能夠與成纖維細胞上的相應受體發生相互作用(圖7d)。
令人驚訝的是,成纖維細胞通過推測的配體-受體相互作用與許多細胞類型相連接。成纖維細胞產生了許多趨化因子(CCL13、CCL19、CCL2、CCL8、CXCL1、CXCL12、CXCL2、CXCL3),與髓樣細胞和T細胞表達的受體相結合,提示成纖維細胞在銀屑病中招募免疫細胞方面起著重要作用(圖7e)。成纖維細胞還產生細胞因子IL-7和IL-15,與T細胞上的IL-7受體和IL-15受體相互作用,促進T細胞增殖和成熟。成纖維細胞通過表達與其受體結合的成纖維細胞生長因子(FGF10、FGF2、FGF7),具有誘導角質細胞過度增殖的能力。我們通過免疫組織化學(IHC)在銀屑病斑塊皮膚中驗證了成纖維細胞生長因子FGF2和FGF7以及它們的受體FGFR2和FGFR3的表達。
髓樣細胞表達多種趨化因子,進一步促進其他白細胞向銀屑病部位的招募(圖7f)。這些趨化因子還與角質細胞和成纖維細胞表達的非典型趨化因子受體(ACKR2、ACKR3、ACKR4)相互作用,引發過度的Th17反應并放大皮膚炎癥。此外,髓樣細胞表達的細胞因子(IL15、IL1A、IL1B、IL6)抑制角質細胞的凋亡。T細胞表達與角質細胞和成纖維細胞表達的非典型趨化因子受體相互作用的細胞因子(CCL3、CCL3L1、CCL4、CCL5、CXCL13)(圖7g)。此外,T細胞表達的IL17A、IL17F和IL22與髓樣細胞和角質細胞表達的受體結合,已知在銀屑病中促進炎癥。
因此,通過整合單細胞轉錄組測序數據、上游調節因子分析、配體-受體結果以及不同細胞類型的空間位置,我們能夠創建一個突出成纖維細胞在銀屑病皮膚中作用的模型。在銀屑病中,SFRP2+成纖維細胞從纖維化狀態轉變為炎癥狀態。這些炎癥性成纖維細胞表達CCL13和CCL19,能夠招募表達相關趨化因子受體的細胞:CCR2+髓樣細胞和CCR7+LAMP3+經典型2型樹突狀細胞A。這些炎癥性成纖維細胞還高水平表達CXCL1,已知能夠招募中性粒細胞,以及CXCL12,已知能夠招募CXCR4+Tc17細胞。CXCL12還可以通過其在表皮上的受體CXCR4作用誘導角質細胞增殖。在表皮中,Tc17分泌的IL-17引發炎癥,通過上皮角質層內的IL-36循環進一步放大,觸發CCL20的釋放,CCL20可以招募CCR6+Tc17以及CXCL1和CXCL8可以招募中性粒細胞,與上皮角質層中的微膿腫一致。
實驗方法
免疫組織化學、scRNA-seq、Seurat、亞型細分、擬時序分析、銀屑病相關遺傳風險變異分析、配體-受體相互作用分析、Spatial-seq、CRISPR/Cas9、qRT-PCR、Seq-Scope樣品處理和分。
參考文獻
Ma, F., Plazyo, O., Billi, A.C. et al. Single cell and spatial sequencing define processes by which keratinocytes and fibroblasts amplify inflammatory responses in psoriasis. Nat Commun 14, 3455 (2023).